Plants are the source of our food, medicine, construction materials, and the foundation of ecosystems. How can we improve the productivity and resilience of plants? What knowledge, technologies, and tools do we need to generate? These are the long-term questions Zhiyong Wang's lab aim to answer in their research.
Plant growth and survival depend on cellular signaling mechanisms through which plant cells monitor and respond to hormonal signals, environmental cues, and internal nutrient status. Brassinosteroid (BR) is a major growth-promoting hormone that effects on plant height, size, and biomass accumulation.
Plant growth and development are also highly sensitive to environmental signals such as light/dark, temperature, and pathogens. Of course, plant growth depends on nutrients including nitrogen and sugars (product of photosynthesis), and nutrient-sensing mechanisms, such as the Target of Rapamycin (TOR) kinase or O-glycosyltransferases (SPINDLY and SECRET AGENT), are essential for viability.
Zhiyong Wang's research dissects the molecular mechanisms underlying growth responses to these internal and external factors, which have major impacts on plant growth and resilience. To achieve a comprehensive and mechanistic understanding of the growth regulatory system, his lab uses broad research approaches and technologies, including genomics, proteomics, chemical proteomics, microscopy, computation, and structural biology.
Accomplishments
The Wang Lab's research has established the framework of molecular networks that explain how nutritional, hormonal, and environmental signals coordinate the cellular decisions of growth, immunity, and acclimation. Most of the former postdocs and students who made these important discoveries are now leading their own labs in academic institutions.
Among the major achievements of our research, Zhiyong Wang's team has illustrated:
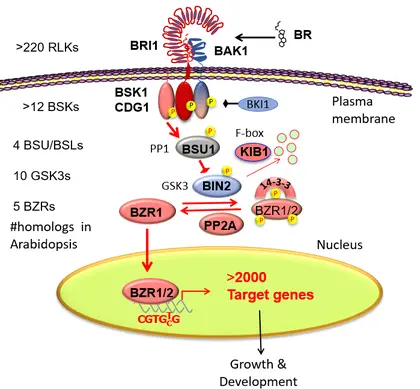
The full brassinosteroid (BR) signaling pathway from the receptor kinase BRI1 to nuclear transcription factor BZR1 and its thousands of target genes.
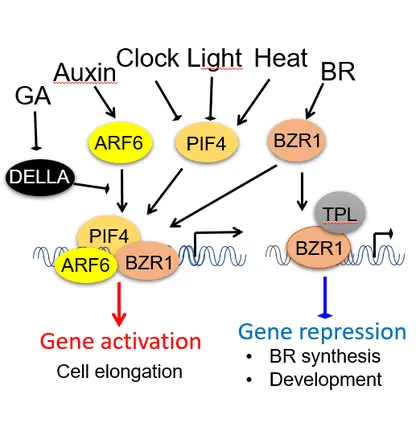
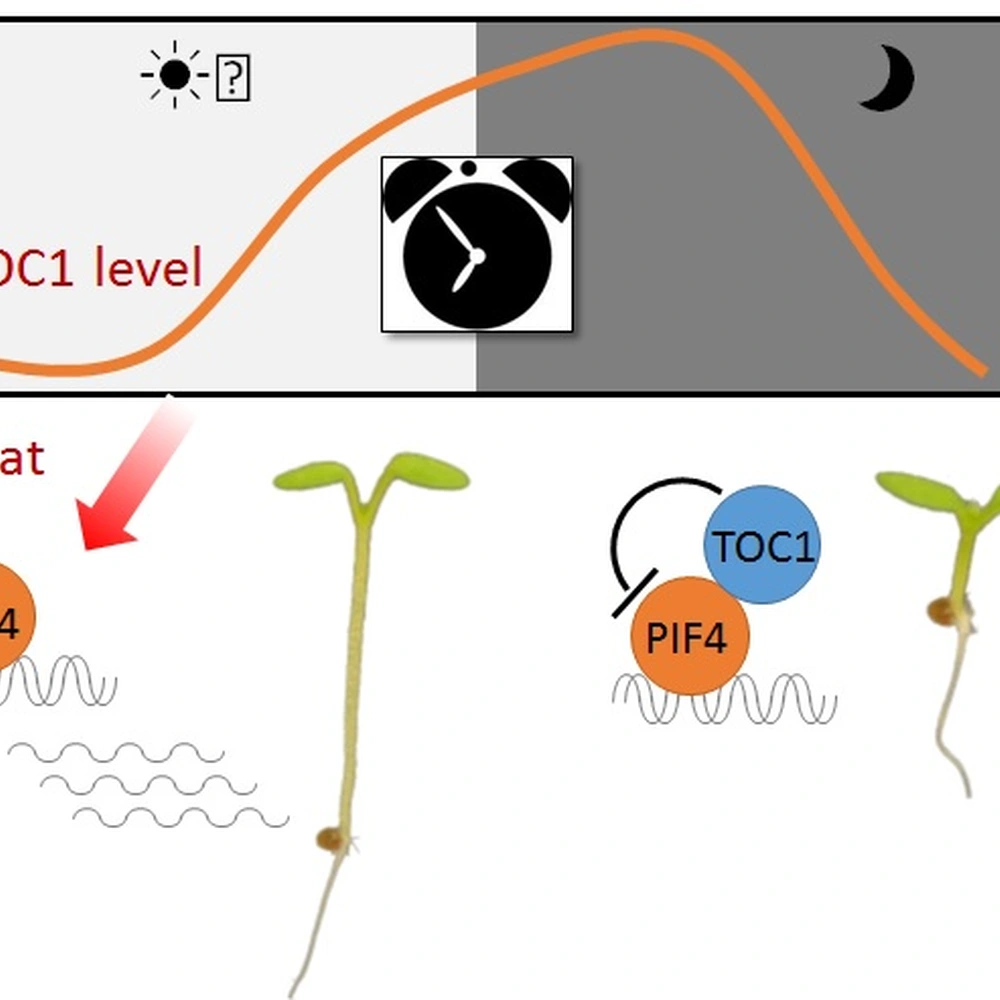
The growth co-regulation by key growth hormones (BR, auxin, gibberellin) and environmental signals (light and temperature) through direct interactions among their responsive transcription factors, a signal integration mechanism named BAP/D module
The spatiotemporal actions of BR in patterning growth and development in the shoot and root tips.
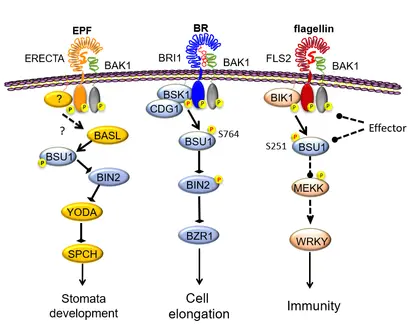
The mechanisms of crosstalk and component-sharing between BR/BRI1 and other receptor kinase pathways that regulate stomata development and immunity.
The expansive BR-response phosphorylation network controlled by the BIN2/GSK3 kinase.
The genetic variations in the BR-response cis-elements contribute to traits in maize.
The expansive nutrient-signaling networks of protein posttranslational modifications by O-linked β-N-acetylglucosamine (O-GlcNAc) and O-fucose.
The significant overlaps between the BR-regulated phosphorylation network and the nutrient-dependent O-glycosylation networks.
Ongoing Work:
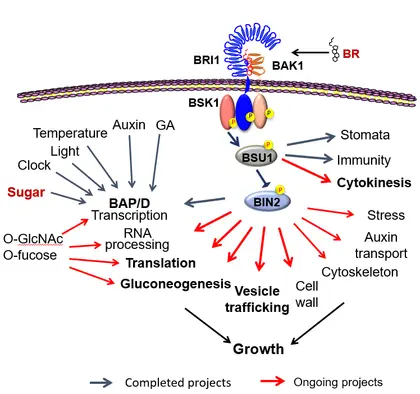
Building upon a large amount of solid data and converging discoveries while taking advantage of the in-house mass spectrometry facility/technologies, our current research continues to make exciting progress toward answering important scientific questions. These include:
How does BR-dependent phosphorylation regulate membrane trafficking, an essential aspect of cell growth?
How do the BR-signaling proteins regulate cytokinesis in plants?
How do cells maintain cell wall integrity during hormone-induced cell expansion?
How do O-GlcNAcylation and O-fucosylation mediate sugar regulation of protein functions and cellular/developmental/physiological processes?
How do BR and sugar signaling, through phosphorylation and O-glycosylation, respectively, co-regulate metabolism and growth?
How do phosphorylation and O-glycosylation crosstalk on common target proteins?
These projects are led by individual postdocs and graduate students, who collaborate and support each other, under my guidance. Together, we are advance a systems-level mechanistic understanding of plant growth and acclimation, and we identify targets and strategies for improving plant productivity and resilience.
Looking Ahead:
What are the main challenges that we still need to overcome? What are the opportunities provided by accumulating knowledge and advancing technologies?
We need to develop tools that enable spatiotemporal manipulation of specific signaling events, and we are developing such tools using nanobodies, molecular sensors, and chemicals/drugs. We would like to expand our research into non-model plants of economic or ecological importance. To do this, we need funding and people to replicate in crops (e.g. maize) some of the productive proteomic experiments (e.g. proximity labeling and O-glycosylation profiling) that we have done in Arabidopsis. We also need to develop better transformation methods to easily transform plants that are difficult or impossible to transform with current methods, and we are testing some novel ideas.
The rapid development in technologies presents exciting opportunities for life science. For example, structures of nearly all proteins can now be predicted by AlphaFold and visualized by cryoEM. This makes it possible to carry out structure-based drug discovery for plant biology. We are using combinations of virtual and experimental screening approaches to identify chemical inhibitors and modulators of plant proteins, developing chemical tools useful for basic research and agricultural application.
Contact Zhiyong Wang
The Wang Lab strives to push the frontier and hope to one day make a real positive impact on our world. If you are interested in joining us, or supporting us, reach out via email to zwang@carnegiescience.edu.
Ethylene plays its essential roles in plant development, growth, and defense responses by controlling the transcriptional reprograming, in which EIN2-C-directed regulation of histone acetylation is the first key step for chromatin to perceive ethylene signaling. But how the nuclear acetyl coenzyme A (acetyl CoA) is produced to ensure the ethylene-mediated histone acetylation is unknown. Here we report that ethylene triggers the accumulation of the pyruvate dehydrogenase complex (PDC) in the nucleus to synthesize nuclear acetyl CoA to regulate ethylene response. PDC is identified as an EIN2-C nuclear partner, and ethylene triggers its nuclear accumulation. Mutations in PDC lead to an ethylene hyposensitivity that results from the reduction of histone acetylation and transcription activation. Enzymatically active nuclear PDC synthesizes nuclear acetyl CoA for EIN2-C-directed histone acetylation and transcription regulation. These findings uncover a mechanism by which PDC-EIN2 converges the mitochondrial enzyme-mediated nuclear acetyl CoA synthesis with epigenetic and transcriptional regulation for plant hormone response.
Ethylene plays its essential roles in plant development, growth, and defense responses by controlling the transcriptional reprograming, in which EIN2-C-directed regulation of histone acetylation is the first key step for chromatin to perceive ethylene signaling. But how the nuclear acetyl coenzyme A (acetyl CoA) is produced to ensure the ethylene-mediated histone acetylation is unknown. Here we report that ethylene triggers the accumulation of the pyruvate dehydrogenase complex (PDC) in the nucleus to synthesize nuclear acetyl CoA to regulate ethylene response. PDC is identified as an EIN2-C nuclear partner, and ethylene triggers its nuclear accumulation. Mutations in PDC lead to an ethylene hyposensitivity that results from the reduction of histone acetylation and transcription activation. Enzymatically active nuclear PDC synthesizes nuclear acetyl CoA for EIN2-C-directed histone acetylation and transcription regulation. These findings uncover a mechanism by which PDC-EIN2 converges the mitochondrial enzyme-mediated nuclear acetyl CoA synthesis with epigenetic and transcriptional regulation for plant hormone response.
Photomorphogenesis is a light-dependent plant growth and development program. As the core regulator of photomorphogenesis, ELONGATED HYPOCOTYL 5 (HY5) is affected by dynamic changes in its transcriptional activity and protein stability; however, little is known about the mediators of these processes. Here, we identified PHOTOREGULATORY PROTEIN KINASE 1 (PPK1), which interacts with and phosphorylates HY5 in Arabidopsis, as one such mediator. The phosphorylation of HY5 by PPK1 is essential to establish high-affinity binding with B-BOX PROTEIN 24 (BBX24) and CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1), which inhibit the transcriptional activity and promote the degradation of HY5, respectively. As such, PPKs regulate not only the binding of HY5 to its target genes under light conditions but also HY5 degradation when plants are transferred from light to dark. Our data identify a PPK-mediated phospho-code on HY5 that integrates the molecular mechanisms underlying the regulation of HY5 to precisely control plant photomorphogenesis.
Photomorphogenesis is a light-dependent plant growth and development program. As the core regulator of photomorphogenesis, ELONGATED HYPOCOTYL 5 (HY5) is affected by dynamic changes in its transcriptional activity and protein stability; however, little is known about the mediators of these processes. Here, we identified PHOTOREGULATORY PROTEIN KINASE 1 (PPK1), which interacts with and phosphorylates HY5 in Arabidopsis, as one such mediator. The phosphorylation of HY5 by PPK1 is essential to establish high-affinity binding with B-BOX PROTEIN 24 (BBX24) and CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1), which inhibit the transcriptional activity and promote the degradation of HY5, respectively. As such, PPKs regulate not only the binding of HY5 to its target genes under light conditions but also HY5 degradation when plants are transferred from light to dark. Our data identify a PPK-mediated phospho-code on HY5 that integrates the molecular mechanisms underlying the regulation of HY5 to precisely control plant photomorphogenesis.
The regeneration of the mammalian skeleton's craniofacial bones necessitates the action of intrinsic and extrinsic inductive factors from multiple cell types, which function hierarchically and temporally to control the differentiation of osteogenic progenitors. Single-cell transcriptomics of developing mouse calvarial suture recently identified a suture mesenchymal progenitor population with previously unappreciated tendon- or ligament-associated gene expression profile. Here, we developed a Mohawk homeobox (Mkx CG ; R26R tdT ) reporter mouse and demonstrated that this reporter identifies an adult calvarial suture resident cell population that gives rise to calvarial osteoblasts and osteocytes during homeostatic conditions. Single-cell RNA sequencing (scRNA-Seq) data reveal that Mkx + suture cells display a progenitor-like phenotype with expression of teno-ligamentous genes. Bone injury with Mkx + cell ablation showed delayed bone healing. Remarkably, Mkx gene played a critical role as an osteo-inhibitory factor in calvarial suture cells, as knockdown or knockout resulted in increased osteogenic differentiation. Localized deletion of Mkx in vivo also resulted in robustly increased calvarial defect repair. We further showed that mechanical stretch dynamically regulates Mkx expression, in turn regulating calvarial cell osteogenesis. Together, we define Mkx + cells within the suture mesenchyme as a progenitor population for adult craniofacial bone repair, and Mkx acts as a mechanoresponsive gene to prevent osteogenic differentiation within the stem cell niche.
The regeneration of the mammalian skeleton's craniofacial bones necessitates the action of intrinsic and extrinsic inductive factors from multiple cell types, which function hierarchically and temporally to control the differentiation of osteogenic progenitors. Single-cell transcriptomics of developing mouse calvarial suture recently identified a suture mesenchymal progenitor population with previously unappreciated tendon- or ligament-associated gene expression profile. Here, we developed a Mohawk homeobox (MkxCG; R26RtdT) reporter mouse and demonstrated that this reporter identifies an adult calvarial suture resident cell population that gives rise to calvarial osteoblasts and osteocytes during homeostatic conditions. Single-cell RNA sequencing (scRNA-Seq) data reveal that Mkx+ suture cells display a progenitor-like phenotype with expression of teno-ligamentous genes. Bone injury with Mkx+ cell ablation showed delayed bone healing. Remarkably, Mkx gene played a critical role as an osteo-inhibitory factor in calvarial suture cells, as knockdown or knockout resulted in increased osteogenic differentiation. Localized deletion of Mkx in vivo also resulted in robustly increased calvarial defect repair. We further showed that mechanical stretch dynamically regulates Mkx expression, in turn regulating calvarial cell osteogenesis. Together, we define Mkx+ cells within the suture mesenchyme as a progenitor population for adult craniofacial bone repair, and Mkx acts as a mechanoresponsive gene to prevent osteogenic differentiation within the stem cell niche.